ヤクシマザル調査マニュアル参考資料
ニホンザルの社会と生態 ver.2023
半谷吾郎
1. ニホンザルはどんなサルか
分類上の位置・形態
ニホンザル(Macaca fuscata)は霊長目オナカザル科マカク属に属します。オナガザル科には他にヒヒ、オナガザル、コロブスなど、非常にたくさんの種類のサルがいて、現在の霊長類の中ではもっとも栄えているグループのひとつだと言えます。これらは全てアジアとアフリカにすんでいます。ニホンザルをはじめとするマカクは約19種あり、北アフリカのモロッコとアルジェリアにすむバーバリマカク一種を除いては、全てアジアにすんでいます。マカクの仲間は地上性が強く、長距離を移動するときは地上を四足で移動する種が大半です。マカクはヒヒと並んで最も適応能力の高いサルです。ニホンザルは、マカクの中でも東南アジアにすむカニクイザル、台湾にすむタイワンザル、インド・ヒマラヤ・中国にすむアカゲザルなどに近い仲間です。これら4種のサルは、マカクの中でも、最後に現れたグループです。適応能力の高いマカクの中でも、とくにたくましいサルたちで、霊長類の分布の中心である熱帯林だけでなく、高山や高緯度地帯、あるいは人家の周辺にも進出し、分布域は広くて生息環境は多様です。
ニホンザルの祖先がいつ日本にわたってきたのかは、化石証拠が少ないためによく分かっていません。40-50万年前に、北京原人の化石と同じ場所で発見された(Macaca robustus)というサルがニホンザルの祖先だといわれています。もっとも古いニホンザルの化石は愛媛県で発掘された20万年前頃のものですから、おおむね30‐50万年前ごろに渡来してきたものと考えてよいと思われます。 ニホンザルは、マカクの中で特に尾が短く、10cmくらいしかありません。ニホンザルは北限のサルですが、これは寒いところの動物は突出部が少なくなるという「アレンの法則」に当てはまります。よくサルというときに長い尾を木の枝にまきつけてぶら下がっている姿を想像する人が多いのですが、そういうことができるのは南アメリカにすむクモザル科のサルだけです(尾を木に巻きつけて、バナナを食べるサルの姿が一般の人の頭の中にあるのは、アメリカのテレビ文化の影響でしょう)。屋久島で売っている土産物に書いてあるサルの絵にも尾が長いものがかなりあります。もう一つ、屋久島にある「動物に注意」という道路標識の看板のサルの絵にも(ついでにシカの絵にも)、重大な間違いがあります。
ニホンザルの分布
ニホンザルがヒトを除く霊長類で最も北にすむことはよく知られています。北は青森県の下北半島から南は屋久島まで、茨城県と長崎県をのぞく43の都府県に生息しています。沖縄と北海道にサルがいたことは一度もありません。屋久島の南から奄美大島までの海は深く、一度も干上がったことがないので、朝鮮半島から渡ってきたニホンザルの祖先は進出できませんでした。津軽海峡は氷河期にしばしば干上がりましたが、当時寒冷のためにニホンザルの分布は関東あたりまで後退しており、そのために北海道には渡る機会がなかったと考えられます。長野県の上高地は、ニホンザルが生息する中で最も寒い地域ですが、ここの平均気温は札幌より低いのです。ですから、今の北海道は決してサルの生息が不可能な場所ではありません。
現在の分布は特に東北地方では局所的で、中部・近畿地方で比較的連続しています。推定頭数は4-5万頭から11万頭と幅があり、実際にはよくわかっていません。1923年当時に比べて、1978年には分布が局所化し、茨城県や種子島など、いくつかの場所ではニホンザルが絶滅しました。一方、2000-2003年に環境省が行った「緑の国勢調査」では、全国を5km四方の区画に分けたときに、生息区画数が20年前より1.5倍に増加しました。現在では、分布は拡大傾向にあるようです。
ヤクシマザルとホンドザル
屋久島のニホンザルはヤクシマザル(Macaca fuscata yakui)といってニホンザルの亜種です。本土のものはホンドザル(Macaca fuscata fuscata)と呼ばれています(調査隊では1996年の調査までは「ヤクザル」という言葉だけを使っていましたが、筆者が霊長類学会で最初に発表したとき、「ヤクザルという言葉はやめてもらいたい。ヤクシマザルが正しい名前である」という指摘を受け、それ以来「ヤクシマザル」を使うようにしています)。50年ほど前には種子島にもニホンザルがいたのですが、絶滅してしまって標本も残っていないために、種子島のサルがホンドザルであったかヤクシマザルであったかは不明です。
もっとも、九州一円で馬小屋に「河童の手」などとしてサルの手が保存されていることがあり、最近種子島でもそのようなものがないか調査が始まりました。もしサルの手が見つかれば、そこから遺伝子を取り出して現在のヤクシマザル・ホンドザルと比較することができるでしょう。 ヤクシマザルは、ホンドザルに比べてからだが小さいこと、毛が長くて粗いこと、アカンボウの毛が黒いことなどが違います。遺伝的には、ホンドザルとヤクシマザルの差はホンドザル同士より10倍以上あります。一方、ヤクザル調査隊も協力したミトコンドリアDNAの変異を調べた研究によると、ヤクシマザルの中の遺伝的多様性は非常に低いことが分かっています。今から7000年前に鹿児島と屋久島のあいだにある現在の硫黄島が大噴火し、屋久島だけでなく西日本一帯に甚大な被害を及ぼしたことが知られており、このときに壊滅的な打撃をこうむったことは間違いありません。
ニホンザルの一生
ニホンザルは生後6ヶ月までをアカンボウと呼び、この期間は全面的に母親に依存しています。その後は母親から離れて同年齢の個体と過ごすことが多くなり、コドモと呼ばれます。早ければオスは5歳頃、メスは4歳頃に性的に成熟し、ワカモノになります。10歳頃に成長が止まり、オトナと呼ばれるようになります。
ニホンザルのメスは一生群れにとどまります。野生状態ではオトナメスは約3年に1回コドモを産みます。一方、オスは性的に成熟したころ生まれた群れを離れ、他の群れに移籍したりヒトリザルになったりします。オスは一つの群れに数年滞在し、次々と移籍を繰り返します。
ニホンザルの寿命は野生状態ではよくわかっていませんが、20歳を越えることは少ないでしょう。メスではコドモを生まなくなってからも数年間生きることがあるようです。餌付け群では、京都嵐山のミノというメスは33歳10ヶ月まで生きました。
2. ニホンザルの社会
メス中心の母系社会
ニホンザルの群れの核になるのはメスの血縁集団です。メスは生涯生まれた群れを出ることはないので、母と娘の親しい関係は一生続きます。母と娘の関係を中心にして姉と妹、おばと姪、祖母と孫娘など、母系の血縁でつながった個体同士が血縁集団を作ります。この血縁集団がいくつも集まったものが群れであると言えます。
一方で、オスは性成熟に達すると群れを出るので、その時点で母親との関係は途絶えます。オスは群れにとっては一時的な滞在者で、数年のうちにまた群れを出て移籍したりヒトリザルになったりします。オスがどんなきっかけで群れを出るかは分かっていませんが、このことによって自分の娘と交尾することが結果的に避けられています。オスは母系社会である群れの「雇われガードマン」だ、という見方もあります。

オスは一生の間に移出入を繰り返す。群れの核は血縁で結ばれたメスとそのコドモで、メスは一生群れに留まる。
順位
ニホンザルの群れでは、オトナオス、オトナメスの中で、明確な順位があります。オスとメスではオスの方が順位が高いことが多いようですが、そうでない場合もあります。 ニホンザルのメスの順位は多くの場合生まれたときに決まり、生涯変わりません。生まれた娘は、母親のすぐ下の順位になります。そのため、上位の家系のコドモは下位の家系のコドモよりも優位になり、妹は姉より優位になります(末子優位)。これは、母親が自分のコドモ、それも一番若いコドモを援助して他の個体を攻撃するうちに、母親がいなくても子供が母親の順位をそっくりコピーするようになるためだと考えられています。ところが、屋久島では姉が妹よりも優位になるようで、末子優位が成立しません。これは、屋久島以外の、末子優位があるとされていたのが餌付け群だからだ、説明されてきました。自然状態で攻撃的交渉がそれほど起こらなければ、母親の援助とは関係なく順位が決まる、というわけです。しかし、同じく餌付けされていない宮城県金華山では、末子優位が見られるらしい、ということが分かってきました。生まれつきの寛容性のような、別の性質による説明が必要かもしれません。 一方、オスは、ふつう最下位で群れに入り、上の順位の個体が移籍して出ていくことで順位を上げていくようです。ですが、中にははじめから一位で群れに入り、群れを乗っ取る個体もいます。筆者が観察していた屋久島H群のムツというオスは、1997年10月までは群れの第5位でしたが、その後離脱し、ヒトリザルとしてすごしていました。翌1998年9月に、群れにいたときとは見違えるほど大きくなって現れ、H群を短期間乗っ取りました。ところが、10月に別のオス、ヤマセに群れをまた乗っ取られ、またヒトリザルになった後、12月に最下位でH群に加入しました。1頭のオスを一生涯追跡し、どのように群れを渡り歩くかを調べるような研究はほとんど行われていませんが、このムツのように劇的な離脱・移籍・乗っ取りを繰り返すことは、それほど珍しいことではないのかもしれません。
当然のことですが、いい食物が優位個体と劣位個体のあいだにあれば、優位個体がそれを取り、劣位のオスは優位なオスの前ではメスと交尾することはできません。ですが、野生状態では食物は分散して分布しているので、優位な個体が独占することは難しいし、発情したメスが他のオスと交尾しないように四六時中見張ることはほとんど不可能です。屋久島のような地形の険しいところでは強いオスから隠れて交尾をすることはそれほど難しいことではありません。また、群れの中に発情したメスが何頭もいるような状況では、順位の高いオスによる独占は難しくなるでしょう。実際に優位な個体がたくさん食物を食べているか、あるいはたくさんコドモを残しているかどうかは、状況次第だと言えるでしょう。
現在では、遺伝子を使ってサルのアカンボウの父親を特定することが可能です。屋久島のニホンザルを対象にした研究によると、たくさん交尾をする優位なオスが必ずしも子どもをたくさん残しているわけではないということが分かってきました。また、メスの場合も、順位の高いメスが一生の間にたくさんの子どもを残すとは限らないようです。
群れサイズを決めるもの-群れの分裂・消滅
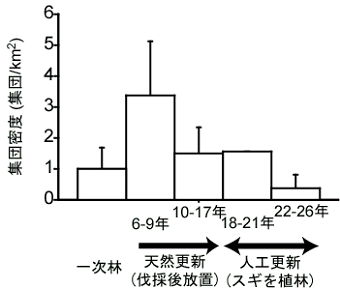
ニホンザルの群れサイズは非積雪地のホンドザルでは50-100頭前後が最も多く、積雪地ではやや小さく最大で70頭前後です。ヤクシマザルはさらに小さく、20-40頭がふつうです。これまで自然群で観察された最大の群れは、餌付け前の大分県高崎山の約160頭、最小が屋久島M群の3頭です。
この3頭になってしまったM群は、最後に残ったメス2頭が隣接群に移籍して消滅しました。群れの消滅は屋久島だけで観察された珍しい事例です。群れが非常に小さくなると、群れの遊動域を他の群れから守れなくなり、最後にはコドモがほとんど産まれなくなって消滅に至るようです。屋久島西部海岸では、観察された限り、群れの消滅が37年間で7例起こっていますが、そのうち5例が、もともと小さかった群れがこのような群れ間競合によってさらに小さくなって消滅に至るというものです。
一方、群れサイズがある程度大きくなると群れが分裂し、サイズが小さくなります。あまり大きい群れだと群れの中で食物をめぐる競合が激しくなり、順位の低い家系の個体が集団で群れを出て行くことで分裂が起こるようです。しかし、餌付けされていると相当群れが大きくなることがあります。例えば高崎山A群は一時1,000頭を越えていました。ここまで大きくなるとサルも群れの中の全部の個体を覚えきれないようです。非常に順位の高いオスが、普段は周縁にいて会うことのない若いオスを見て、群れ外オスと思って逃げ出すことがあるそうです。
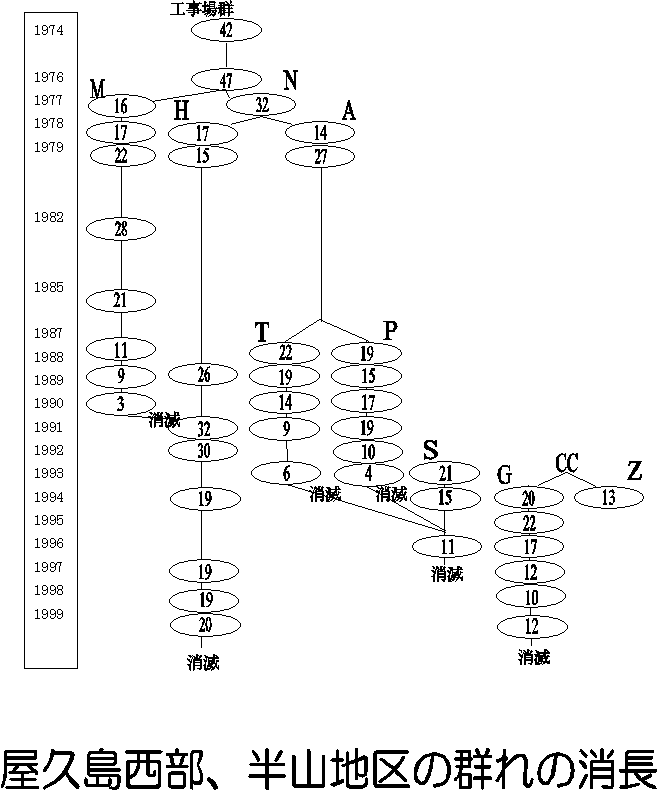
屋久島の西部林道域では、複数の群れを識別した調査が1970年代から継続して行われているので、このような分裂・消滅といった、群れのダイナミックな社会変動が詳しく記録されています。最初に調査された工事場(Ko)群は、1975年当時約50頭で、3回の分裂を経て4群になりましたが、それらは1999年5月までに全て消滅してしまいました。かつての工事場群の遊動域には、南から北から、山の上(東)から侵入してきた様々な群れがいます。
3. ニホンザルの生態
遊動
群れのメンバーは一日中、食べるときも移動するときも休むときも寝るときもおおむね一緒にいます。そして、彼らは決まったねぐらを持ちません。サルのこのような移動様式を「遊動」といいます。
ニホンザルの一日の遊動距離は1kmから3kmがふつうです。群れは遊動域を均等に利用するのではなく、その時期の主要な食物の分布にあわせて遊動します。屋久島の群れの遊動域は1km2以下のことが多く非常に小さいのですが、餌の少ない下北半島の群れでは30 km2もの遊動域を持つことがあります。一頭あたりの遊動域面積は環境の質によって一定で、常緑樹林帯で1.4-1.7ha、落葉樹林帯で8-24haです。また、成熟した針葉樹の人工林は食物としての価値はまったくないため、遊動域内に人工林が含まれていると必然的に遊動域は広くなります。愛知県新城市・鳳来町の市川群は約50km2もありました。
逆に、農作物に依存した群れでは、遊動域は小さくなります。 ニホンザルの群れはメスの血縁集団であると同時に、その土地に結びつきを持った地縁集団でもありますから、群れの遊動域は保守的で、長いこと変わらないでいるようです。しかしときには従来の遊動域を外れて遊動することもあります。1996年秋に、京都/大津の比叡山B群がそのような遊動をしました。この群れは従来の遊動域が5 km2で、35年前にこの群れができたときからほとんど変わっていませんでした。ところが1996年10月末に従来の遊動域を離れて遊動し始め、名神高速道路や三条通を含むいくつかの幹線道路を突っ切って、最大で12km離れたところまで達しました。約一月後にもとの遊動域に戻ったものの、清水寺のすぐそばの大谷本廟や桃山御陵などの観光地に現れたり、山科の市街地を突っ切ったりして、マスコミに大々的に取り上げられ、大騒ぎになりました。筆者はたまたまこの群れを調査していたために対策に駆けずり回らねばならず、大変な思いをしました。
食性
ニホンザルは基本的には植物食性です。日本のように季節変化がはっきりしているところでは、当然のことながら利用できる食物も季節により大きく異なります。春には若い葉、夏にはイチゴの果実や葉、秋には様々な果実や堅果(ドングリ)など、もっとも食物の不足する冬には常緑樹林帯では成熟葉、落葉樹林帯では冬芽や樹皮などを採食します。冬芽や樹皮などは一つ一つが非常に小さかったり、繊維分が多かったりして、日中の70%以上の時間をかけて食べ続けても、まだ必要なエネルギーを確保することができないといわれています。そのため、特に北国のサルは秋に果実や堅果をたくさん食べてそれを脂肪として蓄積し、それによって冬を乗り切ります。また、ニホンザルの出産期は春ですが、これは0歳のアカンボウが最も厳しい冬を最後に迎えることができるようにするためであると考えられます。このように、食物環境・食性の季節変化は、サルの生活に大きな影響を与えています。
動物質も昆虫を中心に利用します。海沿いの群れでは貝など、宮崎県幸島では魚やタコ、屋久島ではトカゲ、カエル、まれにヘビやなども採食するようです。鳥の卵を食べたという報告もあります。同じ霊長類でもチンパンジーなどは肉食が多く、カニバリズム(共食い)さえ知られていますが、野生のニホンザルが哺乳類や鳥の肉を食べたという報告はありません。
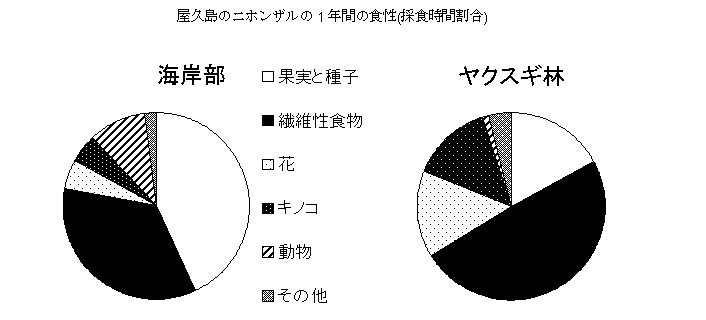
ニホンザルは、環境の変化に対応して柔軟に食性を変化させる能力を持っています。1年間の採食時間割合を屋久島の海岸部とヤクスギ林の群れで比較すると、海岸部では果実や種子が43%を占めるのに対し、ヤクスギ林ではわずか17%に過ぎません。ヤクスギ林ではそのかわりに成熟葉や根などの繊維性の食物が45%を占め、そのほかに花が15%、キノコが14%を占めます。海岸部では繊維性食物は35%にすぎませんし、花やキノコも数%でしかありません。ヤクスギ林の果実生産量は海岸部の約3分の1しかありません。果実はエネルギー含有量が高く繊維が少なくて消化も容易で、ニホンザルが好んで食べる食物です。ヤクスギ林のサルは果実があるときにはたしかに果実をよく食べますが、果実を利用できる時期は限られているため、それ以外のときは繊維性食物をおもに食べるのです。
屋久島海岸部のニホンザルの主要食物
1月 ハドノキ果実、マテバシイ種子、ハナガサノキ葉、ウラジロエノキ葉
2月 マテバシイ種子、ウラジロエノキ葉、ヒメユズリハ葉
3月 バリバリノキ果実、アコウ果実、シャリンバイ種子、ウラジロエノキ葉
4月 イヌビワ果実、シャリンバイ種子、ハゼノキ葉
5月 ヤマモモ果実、アコウ果実
6月 ヤマモモ果実、タブノキ果実
7月 アコウ果実、アカメガシワ種子
8月 クマノミズキ果実、アコウ果実、オオイタビ果実、ハゼノキ種子
9月 アコウ果実、オオイタビ果実、シマサルナシ果実、ハゼノキ種子、マテバシイ種子
10月 アコウ果実、オオイタビ果実、ウバメガシ種子、ハゼノキ種子、マテバシイ種子
11月 シロダモ果実、シマサルナシ果実、シャリンバイ果実、アコウ果実、ハゼノキ種子、ウバメガシ種子
12月 シロダモ果実、シャリンバイ果実、ハマヒサカキ果実、ヒサカキ果実、ハゼノキ種子
屋久島ヤクスギ林のニホンザルの主要食物
1月 ユノミネシダ葉、キノコ、ハイノキ葉、スギ花
2月 ユノミネシダ葉、ハイノキ葉、クロバイ葉、キノコ、スギ花
3月 ヒサカキ花、ヒメノキシノブ葉、ハイノキ葉
4月 ヤブツバキ花、モミ花、キノコ
5月 ヤブツバキ花、モミ花、キノコ、ハイノキ葉
6月 ハイノキ葉、ヤマザクラ果実、ヒサカキ葉
7月 キノコ、ヒサカキ葉、ハイノキ葉、ハイノキ虫こぶ
8月 ハイノキ葉、キノコ、ナナカマド葉、ヒメバライチゴ葉、ハイノキ果実
9月 キノコ、ヤマボウシ種子と果実、ウラジロマタタビ葉、ヒメヒサカキ果実、イスノキ虫こぶ、ハイノキ果実
10月 イスノキ虫こぶと種子、ヒサカキ果実
11月 ヒサカキ果実、イスノキ種子、キノコ、ヤブマオ果実
12月 ハイノキ葉、ヒサカキ果実、キノコ、ヤブマオ果実
サルの数を決めるもの
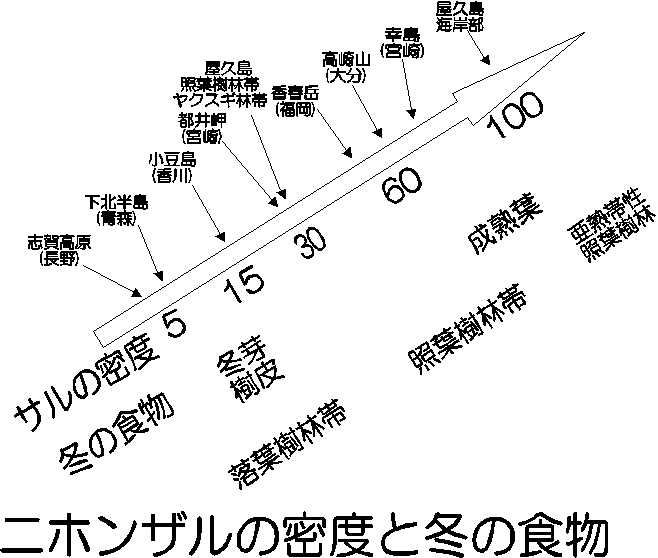
ニホンザルの密度には地域によって大きな違いがあります。狩猟や植生の撹乱など、人為的影響のない環境に限っても、最大で屋久島海岸部の100頭/km2から長野県志賀高原の5頭/km2までの変異があります。ニホンザルの密度は照葉樹林帯と落葉樹林帯で違いがあり、照葉樹林帯は30から70頭/km2、落葉樹林帯は5から15頭/km2のあいだです。屋久島は、われわれヤクザル調査隊の資料によると、海岸部がニホンザルとして最大の100頭/km2、それ以外では30頭/km2前後です。
人為的影響の少ない環境では、サルの数を決めるのは食物資源といってよいでしょう。1997年までのヤクザル調査隊の結果をもとに、屋久島のニホンザルの密度の標高による変異をまとめてみると、密度は海岸部だけが高く、それ以上の標高帯では違いがないことが分かりました。この結果をニホンザルの生息環境と照らしあわせてみると、年間の総果実生産量がこの変化と最も対応しており、1年のうちの果実の利用できない時期の割合や、食物となる樹木の密度には対応していませんでした。また、食物のもっとも不足する冬の食物条件も密度の重要な決定要因です(屋久島はどの標高帯でも冬の食物は常緑樹の成熟葉ですから、屋久島の中での比較では冬の食物条件の違いは無視することができます)。密度の最も低い落葉樹林帯では、サルの冬の食物は冬芽や樹皮です。これらは一つ一つが非常に小さいので、一日中食べ続けても十分な量を採食することができません。それより密度の高い照葉樹林帯では、冬の食物は照葉樹の成熟葉です。成熟葉は果実に比べれば質が劣りますが、一枚一枚が大きいので、とにかく量を稼ぐことができ、冬芽や樹皮よりずっといい食物です。
冬の食物が常緑樹の成熟葉か冬芽や樹皮であるかということと、高質の食物である果実の年間の総生産量と、どちらが密度を決める上で大事なのでしょうか?実はどちらも互いに関係しあっており、どちらがより重要ということはありません。常緑樹の成熟葉にしろ、冬芽や樹皮にしろ、それだけではニホンザルは一日に必要とするエネルギーを賄うことはできません。彼らは秋に果実を食べて蓄積した脂肪を消費することで、冬を乗り切っているのです。同じ脂肪を消費するにしても、常緑樹の成熟葉を食べながらの場合では、冬芽や樹皮を食べながらの場合よりも消費速度が遅いでしょう。つまり脂肪の蓄積、言い換えれば秋の間に1頭あたり食べなくてはいけない食物の量は少なくてすむわけです。このように、脂肪蓄積ということを通じて、冬の食物条件と果実の生産量の両方がニホンザルの密度を決定するのに重要な要因となっているのです。
ニホンザルの個体群動態
生物の数(密度)は、時間とともに変動します。これを個体群動態といいます。ニホンザルの個体群動態にも、いくつかのパターンがあるようです。
宮城県金華山島は、面積10km2あまりの小さな島で、1982年以来、島の全個体数をカウントする調査が継続して行われています。それによると、この島のニホンザルの個体群動態は変動が大きく、1984年と1997年に大量死が見られました。これは、厳冬や、秋のサルの主要食物である何種類かの果実が不作だったことによるものです。金華山のような落葉樹林では、冬には冬芽や樹皮のような食物しか利用できず、それでは必要量を満たすことができません。そのため、秋に果実をしっかり食べ、脂肪として蓄えて冬を乗り切ります。果実が不作で秋にしっかり食べられなかったり、冬が長引いたりすると、蓄えを使い果たす個体が出て、大量死が起こると考えられます。一方、それ以外のふつうの年には、大量死で減った分を取り戻すかのように数が増えていきます。
一方、照葉樹林帯の冬の食物は照葉樹の成熟葉で、これを食べつづけていれば少なくとも死ぬようなことはありません。そうであれば、照葉樹林帯では大量死は起こらないはずです。ところが、1999年冬に、屋久島西部林道域でニホンザルの大量死が起こったことが確認されました。西部林道周辺の半山と呼ばれる地区に分布する5つの群れのうち、二つが半年の間に消滅し、全体としての死亡率が60%にも上りました。前年、1998年の秋がたいへんな不作の年であったのは確かですが、それが唯一の原因であるかどうかはわかりません。西部林道周辺の半山以外の場所では大量死は起こっていないことから、非常に局所的な現象であった可能性が高く、落葉樹林帯での大量死と異なるようです。ちなみに筆者が調査していた群れもこの大量死の時に消滅してしまい、毎日歩けど歩けどサルは見つからず、死体ばかりを見つける辛い日々が1月ほど続きました。
照葉樹林帯では、大量死以外にも、個体数を調節するメカニズムがあるようです。屋久島の西部林道域では、個体数を増やして分裂していく群れがある一方、消滅していく群れもあります。照葉樹林帯では、密度がいずれ飽和状態に達すると、群れと群れのあいだに競合が起こり、小さい群れが消滅することで密度が一定に保たれているのかもしれません。
なお、われわれの調査の大きな目的のひとつは、この個体群動態のメカニズムを屋久島の上部域で明らかにすることにあります。海岸部と同じ常緑樹林ではあるけれど、冬の寒さが厳しく、果実資源に乏しい屋久島上部域では、どのようなメカニズムで個体群密度が維持されているのでしょうか?
森林とサルの関係
サルは食物のほとんどを植物質に依存していますから、サルの生活の様々な側面が植物の影響を受けるのは当然のことです。例えば、サルの出産率は果実の豊凶によって大きく左右されます。秋の果実が豊作であれば、冬を乗り切るための脂肪をたくさん蓄積することができ、翌年の出産率が高くなります。また、サルの毎日の遊動も、どのような植物を利用するかによって決まっています。
これとは逆に、サルのほうが植物に影響を与えることもしばしばあります。霊長類は熱帯で進化した動物ですから、熱帯の植物は霊長類と進化的に長い共存の歴史を持っており、サルに散布されるのに適した形態・味を持った果実をつけるものがたくさんあります。ゴリラだけに種子散布を依存している植物が中央アフリカのガボンで見つかっています。熱帯の植物には、種子散布を霊長類に頼っているものが多くあり、実際、アフリカのウガンダで、サルの密度が多いところと少ないところで動物に散布される植物の実生の密度を比較すると、サルが少ないところでは実生が少ないということが分かりました。熱帯林でサルがいなくなれば、長期的には森林そのものが変質してしまうと考えられます。
他方、温帯の果実は鳥に散布されるように進化してきたと考えられ、熱帯にあるような大きい果実はまず見られません。しかし、屋久島のようにサルの密度が非常に高いところでは、温帯林であってもサルが重要な種子散布の候補者になります。筆者が屋久島のヤクスギ林で調べた結果では、1999年と2000年に鳥が消費した種子数は1.48×106/haと0.58×106/ha、サルが消費した種子数は1.71×106/haと1.34×106/haで、どちらも遜色ないことがわかりました。また、初夏に結実するヤマモモのように、冬鳥がいない場合には、サルのほうがヒヨドリなどの留鳥よりも圧倒的に多くの果実を消費していることが、屋久島海岸部の研究で明らかになっています。興味深いことに、サルが50年前に絶滅した種子島では、その役割を鳥が代替することなく、ほとんどのヤマモモの果実は誰に食べられず落ちてしまうそうです。
ところで、種子散布者として鳥とサルで大きく異なる点があります。鳥は歯がないために果実を丸呑みし、種子は壊されません。ところが、サルは飲み込む前に種子を噛み砕くことがしばしばあります。つまり、サルは種子散布者であると同時に、種子捕食者でもあり、植物にとっては害にもなりうる存在であるということです。植物はそのために非常に堅い種皮を作ったり、種子に有毒物質を含ませたりすることで、噛み割られることを防いでいます。
さらに、サルが植物に一方的に害を与えることもあります。屋久島の西部林道域では、サルの群れが特定のウラジロエノキ、アオモジの葉を繰り返し何度も何度も採食しました。そのため、ウラジロエノキは盆栽状になった木が多く、アオモジは今ではほとんどが枯死してしまいました。金華山ではコブシの木などが同様にサルの採食圧によって枯れ、長野県ではカラマツの木の樹皮をサルがはがして食べ、枯らしてしまったことが報告されています。
他にも、サルが木の上から落とした葉や果実を食べにシカが集まってくるという関係が、屋久島西部海岸と、金華山で報告されています。また、花、キノコ、動物などの採食によっても、植物、菌、動物に影響を及ぼしているはずですが、まだ研究は進んでいません。日本の森林性哺乳類の中で、ニホンザルは体が比較的大きく、集団で生活し、また樹上を利用するなどの特性があります。これらのことからも、森林に対するインパクトは他の哺乳動物とはかなり違ったものかもしれません。